植物分子生理学・植物光生物学
植物分子生理学・植物光生物学
末次 憲之 准教授 (統合生命科学コース) 研究室HP
固着生活を営む植物は、太陽光の光エネルギーを用いて化学エネルギーや有機物を合成する光合成を行い生活しています。植物は光合成を効率よく行うために成長や発生を最適化できるよう、光を光合成のエネルギーとしてだけでなく情報として利用します。植物は多くの“光受容体”を獲得し、光の波長、強さ、方向、照射時間などの情報を統合し、周囲の光環境に適応します。植物は赤色光・遠赤色光受容体フィトクロム、複数の青色光受容体(クリプトクロム、フォトトロピン、FKF1/ZTL/LKP2)、紫外線受容体UVR8を持ち、これらの光受容体の働きにより発芽、成長、発生、成長相転換、概日リズムなど植物の生活環の多くの現象が光によって制御されています。
植物特有な青色光受容体であるフォトトロピン(phototropin、略してphot)は青色光によって活性化する光受容体プロテインキナーゼです。フォトトロピンは光屈性、葉の平滑化、葉緑体運動、気孔開口などを制御し、光合成に必要な光や二酸化炭素利用の効率化を促し、植物体の光合成量そしてバイオマス量の増産を促します(図1)。
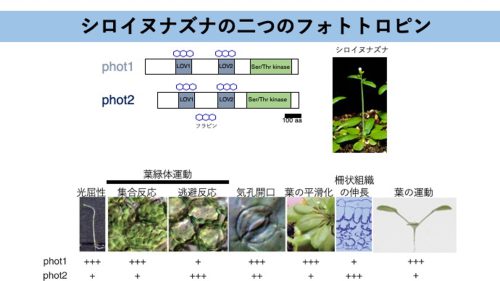
図1.シロイヌナズナの二つのフォトトロピン。
シロイヌナズナはphot1とphot2の二つのフォトトロピン分子種をもつ。どちらも光屈性、葉緑体運動、気孔開口などに冗長的に機能するが、phot1は光屈性や葉緑体集合反応など弱い光で誘導される現象に大きく寄与し、phot2は主に強い光で誘導される反応(葉緑体逃避反応、柵状組織細胞の伸長)を制御する。
私たちの研究室ではフォトトロピンによる光シグナリングを明らかにするために、特に葉緑体運動を研究のモデルとしています(図2)。

図2.葉緑体運動による光受容の最適化。
葉緑体は光合成に必要な光をより多く吸収できるよう弱い光に向かって集まります(集合反応)。集合反応を欠損すると、光の吸収量が減少し光合成量が低下するためバイオマスが減少します。一方、強すぎる光は葉緑体にダメージを与えるので強い光からは逃げます(逃避反応)。逃避反応を欠損すると、葉緑体が強い光のダメージを避けられないため、植物体は白化し枯死します。集合反応はphot1とphot2、逃避反応は主にphot2により制御されています。
シロイヌナズナの葉緑体運動の変異体の分子遺伝学的解析からさまざまな因子が同定されています(図3)。葉緑体運動は細胞骨格のアクチン繊維に依存していますが、他のオルガネラとは異なり、独自のアクチン繊維の重合と運動のメカニズムを利用します(葉緑体[cp]-アクチン繊維)。光受容体フォトトロピンや信号伝達系や運動系で関わる因子のいくつかは、シロイヌナズナだけでなくゼニゴケでも保存されており、陸上植物における葉緑体運動の分子メカニズムの保存性が示唆されていますが、フォトトロピンがどのように信号を発し、その信号がどのように葉緑体に伝わり、どのようにcp-アクチン繊維の重合を制御して葉緑体運動の推進力を生み出すかはわかっていません。私たちの研究室では、植物の光に対する応答反応に興味があり、特に細胞内で葉緑体が光の強さや方向をモニターし、適切な場所に動くための分子メカニズムに興味がある方をお待ちしています。
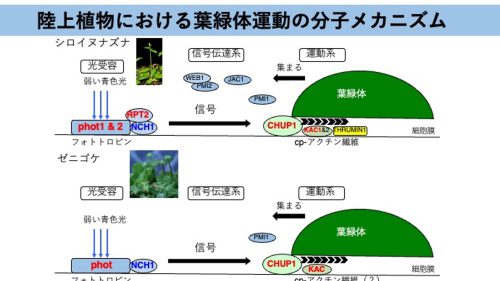
図3.陸上植物における葉緑体運動の分子メカニズム。
葉緑体運動は“光受容”、“信号伝達系”、“運動系”の3つの素過程にわけることができる。陸上植物のモデル植物である種子植物シロイヌナズナとコケ植物ゼニゴケにおいて、フォトトロピンやフォトトロピン結合因子(NCH1/RPT2)、運動系に関わる因子(CHUP1やKAC)が保存されている。