染色体末端の分子生物学から多様な生命現象を紐解く
加納 純子 教授(統合生命科学コース) 研究室HP
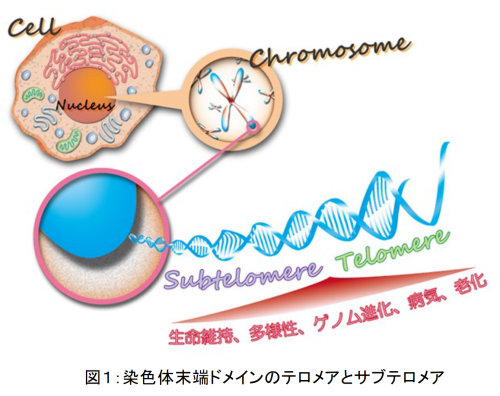
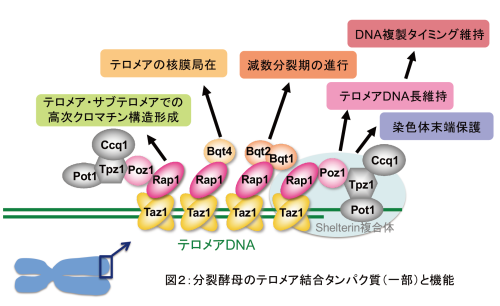
加納研究室では、染色体(遺伝物質であるDNAに様々なタンパク質が結合して形成される構造体)の末端領域であるテロメアやサブテロメアに着目し、それらの分子生物学的解析を行うことにより、様々な生命現象の分子メカニズムを探る研究を行っています。 生物は大きく分けて、真核生物と原核生物に分類されます。多くの原核生物は環状の染色体(核様体とも呼ばれる)を持ちますが、真核生物は線状の染色体を持ちます。線状染色体の最末端部分には、特殊な繰り返し配列のDNAを含むテロメアと呼ばれるドメインが存在し、生命維持に必須の役割を果たすとともに、細胞老化やがん化などと深い関わりがあることが知られています(図1)。我々はこれまでに、テロメアに結合するタンパク質を同定・解析し、様々なテロメア機能を発見してきました(図2:詳細は加納研究室HP参照)。
しかし、テロメアの機能がすべて解明されたわけではありません。そもそもなぜ真核生物は線状の染色体DNAをもつようになったのか? テロメアはどのようにして出現したのか? 真核生物の染色体は原核生物のように環状ではダメなのか? 真核生物には細胞老化という現象が必要なのか?など、当研究室では生命の根本を問う研究も進めています。
テロメアに隣接するドメインはサブテロメアと呼ばれています(図1)。サブテロメアにはテロメアとは異なるDNA配列が見られ、多くの生物で複数の共通ブロック配列が複雑に混在した構造を取っています。このような複雑な構造のために解析が非常に遅れており、染色体の未開の地と言っても過言ではありません。その状況を打開するため、当研究室では世界に先駆けてサブテロメア全破壊株の作製やサブテロメア全DNA配列の決定に成功し、さらに長い間隠されてきたサブテロメアの機能を発見してきました(加納研究室HP参照)。中でも驚いたことに、染色体のセントロメアと呼ばれるドメイン(図1のChromosomeの真ん中の赤い部分)に局在するタンパク質(Sgo2)がサブテロメアにも局在し、セントロメアでの機能とは全く異なる機能を果たすことがわかりました(Tashiro et al., Nature Commun., 2016)。さらに、サブテロメアのDNAは染色体の中で特に変化に富む領域であることもわかりました(Oizumi et al., Nature Commun., 2021)。
サブテロメアの研究は、まだまだやるべきことがたくさんあります。サブテロメアの特徴的な染色体高次構造(クロマチン構造)はどのようにして形成され、どのようにしてサブテロメア領域の範囲が決定されているのか? サブテロメアの変化しやすい性質は何が原因となっているのか? サブテロメアの構造変化が細胞機能にどのような影響をもたらすのか?などは近い将来、解明したい問題です。
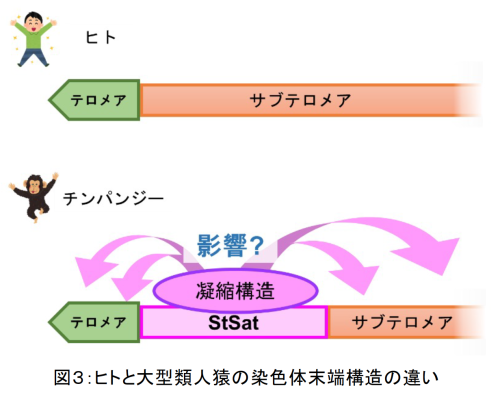
一方、大型類人猿(チンパンジー、ボノボ、ゴリラ、オランウータン)はヒトと進化的に最も近いとされています。特に、ヒトとチンパンジーのDNA配列の違いはわずか数%と言われています。しかし、ヒトとチンパンジーには、骨格、歩行形態、言語の使用、脳の体積、寿命など、様々な違いがあります。このような違いは、わずか数%のDNA配列の違いだけですべて説明できるのでしょうか? 実は、ヒトと大型類人猿の間には、比較ができない染色体構造の根本的な違いが存在しており、ヒトの染色体末端はテロメアーサブテロメアと並んでいますが、大型類人猿ではテロメアとサブテロメアの間にSubtermimal Satellite (StSat)という繰り返し配列が存在します(図3)。このStSat領域が特殊な凝縮構造をとっていることがわかりつつありますが、それが一体どのような機能をもっているのか、それがヒトと大型類人猿の性質の違いに関係しているのか、ヒトをヒトたらしめているものは何か?などを明らかにしようとしています。
生物の根本である遺伝物質DNA。その機能発現を制御する染色体という構造体。染色体は昔から、そしてこれからも研究者の心をつかんで離さない魅力的で神秘的な研究対象です。一緒に生命の神秘のベールを剥いであっと驚く発見をしてみませんか?